Kylling-egg problem ved livets oppstart.
Skrevet av Otangelo Grasso, stoffutvalg herfra.
Livets første molekyl var protein, ikke RNA, antyder ny modell
For forskere som studerer livets opprinnelse, er et av de største kylling-eller--egg-spørsmålene: Hva som kom først-proteiner eller nukleinsyrer som DNA og RNA? (24)
Informasjon kreves for å trekke ut energi. Men Extract Energy er påkrevd for å lagre informasjon. Hva kom først?
Det som endret seg, er at de i stedet for å lukke hullene, har blitt bredere og bredere og naturalistiske forslag mer og mer urealistiske!
I Joseph Hellers klassiske roman om andre verdenskrig, Catch-22, kan en luftfartøy unnskyldes fra kampplikt for å være gal. Men en regel spesifiserte at han først måtte be om en unnskyldning, og alle som ba om en unnskyldning fra kampplikt var åpenbart ikke gal, så slike forespørsler ble alltid nektet. Regelen som gjorde det umulig å bli unnskyldt fra kampplikt ble kalt "Catch-22."
Bilde 1. Motsridende problemer -som ikke lar seg løse samtidig
 Oksygen og liv har et Catch22 forhold
Oksygen og liv har et Catch22 forhold
Catch 22 er en situasjon der en handling har konsekvenser som gjør det umulig å forfølge handlingen. Oksygen er veldig skadelig for livet. Samtidig er oksygen nødvendig for å tilveiebringe ozonlaget som beskytter livet mot ultrafiolett stråling (UVR) som kommer fra solen. Hvis cyanobakterier kom før oksygen, fordi det er årsaken til oksygen, ville cyanobakterier ha måttet utvikle flere former for beskyttelse for å dempe skaden fra UVR: unngåelse, rensing, screening, reparasjon og programmert celledød. UVR -skader er imidlertid øyeblikkelig, og tiden som trengs for å "utvikle" beskyttelse mot den via naturlig seleksjon, utrolig treg. Så UVR -skader ville oppstå før slike forsvarsmekanismer kunne utvikle seg. (3)
Jern-svovelklynger
Svovel er et essensielt element, og er en bestanddel av mange proteiner og kofaktorer. Jern-svovel (FES) sentre er essensielle proteinkofaktorer i alle former for liv. Ulike biosyntetiske veier ble funnet å være tett sammenkoblet gjennom komplekse krysstale-mekanismer som avgjørende avhenger av biotilgjengelig-heten til metallionene jern, molybden, wolfram, nikkel, kobber og sink. Proteiner som krever Fe/S-klynger på sitt aktive sted har blitt lokalisert i mitokondrier, cytosol og kjerner der de er involvert i ganske forskjellige funksjoner som TCA-syklus, aminosyrebiosyntese, ribosom og mitokondriell respirasjon, samfaktorbiosyntese, ribosomsamling, regulering av proteinoversettelse, DNA -replikasjon og DNA -reparasjon. Derfor er prosessen med jern-svovelbiosyntese avgjørende for nesten alle livsformer. Forekomsten av disse proteinene på de metabolske traséene til de fleste organismer fører til at noen forskere teoretiserer at jern-svovelforbindelser hadde en betydelig rolle i livets opprinnelse i jernsulfur verdensteorien. Jern-svovel-verdenshypotesen er et sett med forslag for livets opprinnelse og den tidlige utviklingen av livet avansert i en serie artikler mellom 1988 og 1992 av Günter Wächtershäuser. FES-klyngemontering er en kompleks prosess som involverer mobilisering av FE- og S-atomer fra lagringskilder, deres montering til [FE-S] form, deres transport til spesifikke cellulære lokasjoner og deres overføring til mottaker-apoproteiner. NAR1 er en essensiell komponent i et cytosolisk Fe/S -proteinmonteringsmaskineri. Kreves for modning av ekstramitokondrielle Fe/S -proteiner. (12) Dermed er NAR1 både et mål og en komponent i det cellulære FE/S -proteinbiogenesemaskineriet som skaper en interessant "kylling- og egg" -situasjon for sin modningsprosess konserverte jern -svovel (Fe - S) klynger finnes i en voksende familie av familie av Metalloproteiner som er involvert i prokaryote og eukaryotisk DNA -replikasjon og reparasjon. (20)
Derfor måtte de eksistere før livet begynte, siden DNA -replikasjonsenzymer og proteiner avhenger av dem. De krever imidlertid også komplekse proteiner og enzymer for å bli syntetisert. Det er et klassisk kylling/eggproblem.
Ribonukleotidreduktase
Dette er en av de mest essensielle enzymene i livet. Ribonukleotidreduksjon er den eneste veien for de novo -syntese av deoksyribonukleotider i eksisterende organismer. Denne kjemisk krevende reaksjonen katalyseres av ribonukleotidreduktase (RNR). Mekanismen er ansett som usannsynlig å bli katalysert av et ribozym, og skapte et gåte om hvordan byggesteinene for DNA ble syntetisert ved overgangen fra RNA til DNA-kodede genomer. Biosyntese DNA er laget av RNA. Deoksynukleotidene er laget av nukleotider med ribonukleotidreduktaser (RNR), og produserer uracil-DNA eller U-DNA. Uracil blir deretter omdannet til tymin ved å tilsette en metylgruppe, lage tymin-DNA eller T-DNA, den typen som faktisk brukes. Reaksjonen katalysert av RNR er strengt bevart i alle levende organismer. Videre spiller RNR en kritisk rolle i å regulere den totale frekvensen av DNA -syntese, slik at DNA til cellemasse opprettholdes i et konstant forhold under celledeling og DNA -reparasjon. Et noe uvanlig trekk ved RNR -enzymet er at det katalyserer en reaksjon som fortsetter via en frie radikal virkningsmekanisme. Substratene for RNR er ADP, BNP, CDP og UDP. DTDP (deoksytymidindifosfat) syntetiseres av et annet enzym (tymidylatkinase) fra DTMP (deoksytymidinmonofosfat). Det jernavhengige enzymet, ribonukleotidreduktase (RNR), er essensielt for DNA-syntese. RNR gir en essensiell kobling mellom RNA og DNA -verden. Det bringer oss til den klassiske kylling og egg, Catch22 -situasjonen. RNR -enzymer er påkrevd for å lage DNA. DNA er imidlertid pålagt å lage RNR -enzymer. Hva kom først ?? Vi kan med høy sikkerhet konkludere med at dette enzymet begraver alle RNA -verdensfantasier, og enhver mulighet for overgang fra RNA til DNA -verdensscenarier. (21)
Bilde 2. Gjensidig avhengighet - samtidig forekomst
Tymin
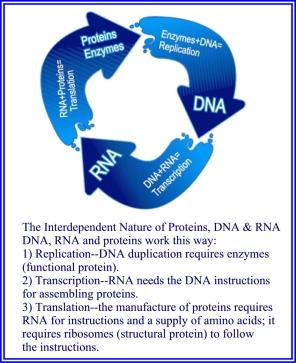 DNA kan bare replikeres i nærvær av spesifikke enzymer (beskrevet nedenfor) som bare kan produseres av det allerede eksisterende DNA. Hver er helt essensiell for den andre, og begge må være til stede for at DNAet kan formere seg. Derfor må DNA ha eksistert i begynnelsen for at livet skal kontrolleres av DNA. Scott M. Huse, "The Collapse of Evolution", Baker Book House: Grand Rapids (Michigan), 1983 P: 93-94
DNA kan bare replikeres i nærvær av spesifikke enzymer (beskrevet nedenfor) som bare kan produseres av det allerede eksisterende DNA. Hver er helt essensiell for den andre, og begge må være til stede for at DNAet kan formere seg. Derfor må DNA ha eksistert i begynnelsen for at livet skal kontrolleres av DNA. Scott M. Huse, "The Collapse of Evolution", Baker Book House: Grand Rapids (Michigan), 1983 P: 93-94
Tymidylatsyntaser (Thy) er viktige enzymer i syntesen av deoksytymidylat, 1 av de 4 byggesteinene til DNA. Som sådan er de essensielle for alle DNA-baserte livsformer og impliseres derfor i den antatte overgangen fra RNA-genomer til DNA-genomer. To ikke -relaterte dine enzymer, Thya og Thyx, er kjent for å katalysere den samme biokjemiske reaksjonen.
Tymidylatsyntase (Thy) er et grunnleggende enzym i DNA-syntese fordi det katalyserer dannelsen av deoksytymidin 5'-monofosfat (DTMP) fra deoksyuridin 5'-monofosfat (dump). I flere tiår var det bare en familie av tymidylatsyntase-enzymer, og dens tilstedeværelse ble ansett som nødvendig for å opprettholde alle DNA-baserte livsformer. Deretter ble et gen som koder for et alternativt enzym oppdaget og karakterisert, og det nye enzymet ble kalt Thyx, mens det andre enzymet ble omdøpt til. Selv om begge reaksjonene oppnår samme nøkkeltrinn, er reaksjonsmekanismene eller trinnene, katalysert av FDT -ene og TS -enzymer strukturelt forskjellige. De to enzymene, Thya og Thyx, ble funnet å ha utpreget forskjellige sekvenser og strukturer, og dermed henvist til uavhengig opprinnelse. (21)
Det er interessant, ettersom vi finner to distinkte enzymer med to forskjellige sekvenser og strukturer som syntetiserer den samme reaksjonen, og dermed er et eksempel på konvergens helt i begynnelsen. Hvor fjernkontroll var sjansen for at dette skulle skje med naturlige midler, med tanke på at konvergens ikke favoriserer naturalistiske forklaringer?
Som Stephen J.Gould skrev: “… Ingen finale kan spesifiseres i starten, ville ingen noen gang skje en gang på samme måte, fordi noen vei fortsetter gjennom tusenvis av usannsynlige stadier. Endre enhver tidlig hendelse, aldri så lett, og uten tilsynelatende betydning på den tiden, og evolusjonskaskader til en radikalt annerledes kanal. (1)
Stephen J. Gould, Wonderful Life: The Burgess Shale and the Nature of History (New York, NY: W.W. Norton & Company, 1989), 51.
I kraft av deres funksjon og filetisk fordeling er Thys eldgamle enzymer, noe tilstedeværelse av et gen som koder for ditt i genomet til de vanlige forfedrene til eukaryoter, bakterier og archaea. Dermed kan det å spore tilbake veien for gener som koder for Thya og Thyx, belyset det aktivt diskuterte bredere spørsmålet om opprinnelsen til viral og cellulær DNA
Dette bringer oss til det samme problemet som med ribonukleotidreduktase-enzymer (RNR), som er den klassiske kylling- og egg, Catch22 -situasjonen. Thya og Thyx -enzymer er pålagt å lage DNA. DNA er imidlertid pålagt å lage disse enzymene. Hva kom først ?? Vi kan med høy sikkerhet konkludere med at dette enzymet begraver alle RNA -verdensfantasier, og enhver mulighet for overgang fra RNA til DNA -verdensscenarier, siden begge måtte komme til å eksistere samtidig.
Hva kom først, proteiner eller proteinsyntese?
 Både transkripsjons- og oversettelsessystemene er avhengige av mange proteiner, hvorav mange er i fellesskap nødvendige for at proteinsyntese i det hele tatt skal oppstå. Likevel er alle disse proteinene laget av denne prosessen. Proteiner involvert i transkripsjon som RNA -polymeraser, for eksempel, er bygget av instruksjoner som er ført på et RNA -transkripsjon. Oversettelse av RNA -transkriptet avhenger av andre spesialiserte enzymer som syntetaser, men informasjonen for å bygge disse enzymene er oversatt under oversettelsesprosessen som syntetaser seg selv tilrettelegges. Biokjemiker David Goodsell beskriver problemet, "Den viktigste molekylære prosessen som gjør det moderne livet mulig er proteinsyntese, siden proteiner brukes i nesten alle aspekter av levende. Syntese av proteiner krever en tett integrert reaksjonssekvens, hvorav de fleste er utført selv av proteiner. " Eller som Jacques Monod bemerket i 1971: "Koden er meningsløs med mindre den er oversatt. Den moderne cellens oversettelsesmaskiner består av minst femti makromolekylære komponenter som selv er kodet i DNA: koden kan ikke oversettes på annen måte enn av Products of Translation." (Forskere vet nå at oversettelse faktisk krever mer enn hundre proteiner.) Den integrerte kompleksiteten i cellens informasjonsbehandlingssystem har ført til en viss dyp refleksjon. Som Lewontin spør: "Hva gjør proteinene som er nødvendige for å lage proteinet?" Som David Goodsell uttrykker det, er dette "en av de ubesvarte gåtene i biokjemi: Hva som kom først, proteiner eller proteinsyntese? Hvis proteiner er nødvendig for å lage proteiner, hvordan kom det hele i gang?" Sluttresultatet av proteinsyntese er nødvendig før det kan begynne.
Både transkripsjons- og oversettelsessystemene er avhengige av mange proteiner, hvorav mange er i fellesskap nødvendige for at proteinsyntese i det hele tatt skal oppstå. Likevel er alle disse proteinene laget av denne prosessen. Proteiner involvert i transkripsjon som RNA -polymeraser, for eksempel, er bygget av instruksjoner som er ført på et RNA -transkripsjon. Oversettelse av RNA -transkriptet avhenger av andre spesialiserte enzymer som syntetaser, men informasjonen for å bygge disse enzymene er oversatt under oversettelsesprosessen som syntetaser seg selv tilrettelegges. Biokjemiker David Goodsell beskriver problemet, "Den viktigste molekylære prosessen som gjør det moderne livet mulig er proteinsyntese, siden proteiner brukes i nesten alle aspekter av levende. Syntese av proteiner krever en tett integrert reaksjonssekvens, hvorav de fleste er utført selv av proteiner. " Eller som Jacques Monod bemerket i 1971: "Koden er meningsløs med mindre den er oversatt. Den moderne cellens oversettelsesmaskiner består av minst femti makromolekylære komponenter som selv er kodet i DNA: koden kan ikke oversettes på annen måte enn av Products of Translation." (Forskere vet nå at oversettelse faktisk krever mer enn hundre proteiner.) Den integrerte kompleksiteten i cellens informasjonsbehandlingssystem har ført til en viss dyp refleksjon. Som Lewontin spør: "Hva gjør proteinene som er nødvendige for å lage proteinet?" Som David Goodsell uttrykker det, er dette "en av de ubesvarte gåtene i biokjemi: Hva som kom først, proteiner eller proteinsyntese? Hvis proteiner er nødvendig for å lage proteiner, hvordan kom det hele i gang?" Sluttresultatet av proteinsyntese er nødvendig før det kan begynne.
Bilde 3. Protein-chaperoner oppfyller maskinkrav
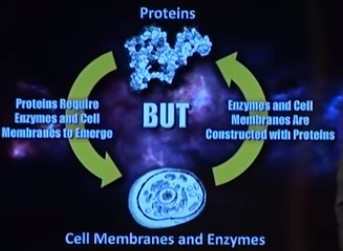
Gjensidig avhengighet av lipidmembraner og membranproteiner
Cellemembranen inneholder forskjellige typer proteiner, inkludert ionekanalproteiner, protonpumper, G -proteiner og enzymer. Disse membranproteinene fungerer samarbeidsvillig for å la ioner trenge gjennom lipid -dobbeltlaget. Den gjensidige avhengigheten av lipidmembraner og membranproteiner antyder at lipid-dobbeltlag og membranproteiner ko-utviklet seg sammen med membranbioenergetikk. Tullet i denne påstanden er tydelig. Hvordan kunne membranproteinene utvikle seg, hvis de måtte produseres i maskineriet, beskyttet av cellemembranen? Cellemembranen inneholder forskjellige typer proteiner, inkludert ionekanalproteiner, protonpumper, G -proteiner og enzymer. Disse membranproteinene fungerer samarbeidsvillig for å la ioner trenge gjennom lipid -dobbeltlaget. ER- og Golgi -apparatet utgjør sammen endomembranrommet i cytoplasmaet til eukaryote celler. Endomembrane-rommet er et viktig sted for lipidsyntese, og ER er der ikke bare lipider blir syntetisert, men membranbundne proteiner og sekretoriske proteiner også blir laget.
Så for å lage cellemembraner, er det endoplasmatiske retikulum påkrevd. Men også Golgi -apparatet, peroksisomet og mitokondriene. Men disse fungerer bare, hvis de er beskyttet og innkapslet i cellemembranen. Hva kom først, cellemembranen, eller endoplasmatisk retikulum? Dette er en av mange andre Catch22 -situasjoner i cellen, som indikerer at cellen ikke kunne dukke opp på en trinnvis gradvis måte, slik talsmenn for naturlige mekanismer ønsker å få oss til å tro.
 Ikke bare er cellemembranen intrikate og komplekse (og absolutt ikke tilfeldige), men den har innstillingsparametere for i hvilken grad fosfolipidhalene er mettede. Det er et annet eksempel på en sofistikert biologisk design som evolusjonister bare kan spekulere i. Tilfeldige mutasjoner må ha heldigvis samlet molekylære mekanismer som føler miljøutfordringer og reagerer på dem ved å endre fosfolipidpopulasjonen i membranen på akkurat den rette måten. Slike design er enormt nyttige, så selvfølgelig ville de blitt bevart av naturlig seleksjon. Det er nok et eksempel på hvor dum evolusjonsteori er i lys av vitenskapelige fakta. (16)
Ikke bare er cellemembranen intrikate og komplekse (og absolutt ikke tilfeldige), men den har innstillingsparametere for i hvilken grad fosfolipidhalene er mettede. Det er et annet eksempel på en sofistikert biologisk design som evolusjonister bare kan spekulere i. Tilfeldige mutasjoner må ha heldigvis samlet molekylære mekanismer som føler miljøutfordringer og reagerer på dem ved å endre fosfolipidpopulasjonen i membranen på akkurat den rette måten. Slike design er enormt nyttige, så selvfølgelig ville de blitt bevart av naturlig seleksjon. Det er nok et eksempel på hvor dum evolusjonsteori er i lys av vitenskapelige fakta. (16)
Bilde 4. Cellemembraner er intrikate og komplekse
DNA - enzymsystemet er irreducibelt sammensatt
Et ofte uopptatt aspekt av kompleksitet er hvordan tRNA blir tildelt de riktige aminosyrene. For at DNA -språket skal oversettes riktig, må hvert tRNA -kodon festes til riktig aminosyre. Hvis dette avgjørende trinnet i DNA -replikasjon ikke er funksjonelt, brytes DNA -språket ned. Spesielle enzymer kalt aminoacyl - tRNA -syntetaser (AARS) sikrer at riktig aminosyre er festet til et tRNA med riktig kodon gjennom en kjemisk reaksjon kalt "aminoacylering." Nøyaktig oversettelse krever ikke bare at hvert tRNA tildeles riktig aminosyre, men også at det ikke blir aminoacylert av noen av AARS -molekylene for de andre 19 aminosyrene. én biokjemi lærebok bemerker at fordi alle AARS -er katalyserer lignende reaksjoner på forskjellige lignende tRNA -molekyler, trodde man at de "utviklet seg fra en felles stamfar og derfor skulle være strukturelt relatert." (Voet og Voet s. 971-975) Imidlertid var dette ikke tilfelle, da "AARSS danner en mangfoldig gruppe på [over 100] enzymer ... og det er liten sekvenslikhet blant syntetaser som er spesifikke for forskjellige aminosyrer." (Voet og Voet s. 971-975) Utrolig nok er disse AARS-ene selv kodet for av DNA: Dette danner essensen av et kylling-egg-problem. Enzymene selv bygger hjelp til å utføre selve oppgaven som konstruerer dem! 9
Hvilket kom først, glykolyse for å lage energi eller energi fra glykolyse som er nødvendig for å lage enzymer? Uten enzymene kunne ikke glykolyse oppstå for å produsere ATP. Men uten ATP kunne ikke disse enzymene produseres. (10)
Et enklere opprinnelse for livet
DNA-replikasjon kan ikke fortsette uten hjelp fra en rekke proteiner-medlemmer av en familie med store molekyler som er kjemisk veldig forskjellige fra DNA. Proteiner, som DNA, er konstruert ved å koble underenheter, aminosyrer i dette tilfellet, sammen for å danne en lang kjede. Celler bruker tjue av disse byggesteinene i proteinene de lager, og gir en rekke produkter som er i stand til å utføre mange forskjellige oppgaver-proteiner er håndymen til den levende cellen. Deres mest berømte underklasse, enzymer, fungerer som ekspedisjoner, og fremskynder kjemiske prosesser som ellers vil finne sted for sakte til å være til nytte for livet. Ovennevnte beretning får tankene til den gamle gåten: Hva som kom først, kyllingen eller egget? DNA har oppskriften på proteinkonstruksjon. Likevel kan ikke denne informasjonen hentes eller kopieres uten hjelp fra proteiner. Hvilket stort molekyl dukket da opp først med å komme i gang-proteiner (kyllingen) eller DNA (egget)? (11)
mRNA er nødvendig for å danne kjerne porekompleks. Men uten kjerne- porekomplekset kan ikke mRNA utarbeides for oversettelse i ribosomet. Det er en Catch22-situasjon .. (1)
---
 Kylling og egg
Kylling og egg
Yarus modell reiser også et betydelig kylling-og-egg-paradoks. Meyer og Nelson forklarer: Fordi de biosyntetiske traséene involverer mange enzymer, vil eksisterende celler kreve et eksisterende translasjonssystem for å lage dem. Siden forsøk på å forklare opprinnelsen til den genetiske koden også prøver å forklare opprinnelsen til oversettelsessystemet (det kan faktisk ikke være noen oversettelse uten kode), gir Yarus et al. Funn et akutt kylling- og eggproblem. Hva som kom først, aptamer-aminosyresaffiniteter som Yarus et al. foreslo som grunnlag for kode- og oversettelsessystemet, eller oversettelsessystemet som ville vært nødvendig for å produsere disse aminosyrene (og dermed aptamer-aminosyresaffiniteter) i utgangspunktet? (2)
Et nytt kylling-og-egg-paradoks relatert til livets opprinnelse (12)
Bilde 5. Motstridende krav -som begge må oppfylles
Celler kunne ikke ha utviklet seg uten virus, da de trenger omvendt transkriptase (som bare finnes i virus) for å bevege seg fra RNA til DNA. Med andre ord, i stedet for å bidra til å løse problemet med livets opprinnelse på jorden, har nyere forskning bare tjent til å fremheve et av de sentrale paradoksene. Og likevel rapporterer Science Media de siste funnene som om løsningen er rett rundt hjørnet. Finner du ikke det bare litt rart? "For å gå fra RNA til DNA, trenger du et enzym som kalles omvendt transkriptase," sa Dolja. "Det finnes bare i virus som HIV, ikke i celler. Så hvordan kunne celler begynne å bruke DNA uten hjelp av et virus? ” (1,3)
Proteiner er påkrevd for å lage proteiner (14)
Trusselen om autodestruksjon stammer fra den sirkulære naturen til proteinsyntese. Proteiner utgjør mange komponenter i cellens proteinproduksjons-maskineri. Med andre ord bruker cellen proteiner for å lage proteiner. Så hvis proteinproduksjons-maskineriet ble samlet med mangelfulle deler, ville cellen ikke klarer å produsere proteiner nøyaktig. En slik produksjonssvikt vil føre til at proteinproduksjonssystemer blir stadig mer utsatt for hver påfølgende runde med proteinsyntese. Proteinproduksjonssystemer som består av mangelfulle komponenter vil være mer sannsynlig å produsere mangelfulle proteiner. Denne kjedereaksjonen ville kaskade ut av kontroll og ganske raskt føre til cellenes selvdestruksjon. Effektive kvalitetssikrings-prosedyrer må være på plass for proteinproduksjon eller liv vil ikke være mulig.
Oppretting av dobbeltstrenget DNA involverer også strengoverføring, der det er en translokasjon av kort DNA-produkt fra initial RNA-avhengig DNA-syntese til akseptormalregioner i den andre enden av genomet, som senere nås og behandles av den revers transkriptase for sin DNA-avhengige DNA-aktivitet
Prosessen med DNA -replikasjon avhenger av mange separate proteinkatalysatorer for å slappe av, stabilisere, kopiere, redigere og spole tilbake den originale DNA -meldingen. I prokaryote celler involverer DNA -replikasjon mer enn tretti spesialiserte proteiner for å utføre oppgaver som er nødvendige for å bygge og nøyaktig kopiere det genetiske molekylet. Disse spesialiserte proteinene inkluderer DNA-polymeraser, primaser, helikaser, topoisomeraser, DNA-bindende proteiner, DNA-ligaser og redigering enzymer.(38) DNA trenger disse proteinene for å kopiere den genetiske informasjonen inneholdt i DNA. Men proteinene som kopierer den genetiske informasjonen i DNA er selv bygget av den informasjonen. Dette utgjør igjen det som i det minste er en gåte: Produksjon av proteiner krever DNA, men produksjonen av DNA krever proteiner.
 "Catch-22" i livets opprinnelse er dette: DNA kan gjenskape, men det trenger enzymer for å katalysere prosessen. Proteiner kan katalysere DNA -dannelse, men de trenger DNA for å spesifisere riktig sekvens av aminosyrer. ” Faktisk er opprinnelsen til den genetiske koden en ond sirkel: proteindaskiner er nødvendig for å lese DNA, men disse proteinmaskinene er i seg selv kodet på DNA. Videre bruker de energi, som krever ATP, laget av nano-motorisk ATP-syntase. Likevel er dette kodet på DNA, dekodet av maskiner som trenger ATP. Proteinene er maskineriet, og DNA er reproduktivmaterialet, men begge er nødvendige samtidig for at cellen i det hele tatt skal fungere. Og selvfølgelig ville dette være ubrukelig uten informasjon å reprodusere.
"Catch-22" i livets opprinnelse er dette: DNA kan gjenskape, men det trenger enzymer for å katalysere prosessen. Proteiner kan katalysere DNA -dannelse, men de trenger DNA for å spesifisere riktig sekvens av aminosyrer. ” Faktisk er opprinnelsen til den genetiske koden en ond sirkel: proteindaskiner er nødvendig for å lese DNA, men disse proteinmaskinene er i seg selv kodet på DNA. Videre bruker de energi, som krever ATP, laget av nano-motorisk ATP-syntase. Likevel er dette kodet på DNA, dekodet av maskiner som trenger ATP. Proteinene er maskineriet, og DNA er reproduktivmaterialet, men begge er nødvendige samtidig for at cellen i det hele tatt skal fungere. Og selvfølgelig ville dette være ubrukelig uten informasjon å reprodusere.
NAR1 er både et mål og en komponent i det cellulære FE/S -proteinbiogenesemaskineriet som skaper en interessant "kylling- og egg" -situasjon for sin modningsprosess (Balk et al., 2004). 5
Hvordan i all verden kan korrekturlesende enzymer dukke opp, spesielt med denne graden av troskap, når de er avhengige av selve informasjonen de er designet for å beskytte? Tenk på det. Dette er et Catch-22 for darwinister. (6)
Bilde 6. Det er ikke 'galt' å spørre
Koonin, The Logic of Chance, Side 231:
Så en uunngåelig (selv om kanskje motintuitiv) konklusjon fra den komparative analysen av eldgamle paralogøst sammenheng mellom proteinkomponenter i oversettelsessystemet er at med det interessante unntaket av kjerne ribosomale proteiner, er alle proteiner som spiller viktige roller i moderne oversettelse produkter av produkter av produkter En lang og kompleks utvikling av forskjellige proteindomener. Her kommer Catch-22: For at all denne proteinutviklingen skal oppstå, kreves et nøyaktig og effektivt oversettelsessystem. Dette primordiale oversettelsessystemet trenger kanskje ikke å være så bra som den moderne versjonen, men det virker som om en sikker innsats, som må ha vært innenfor en størrelsesorden fra den moderne når det gjelder troskap og oversettelseshastigheter for å gjøre proteinutvikling mulig. Fra alt vi vet om det moderne oversettelsessystemet, er imidlertid dette presisjonsnivået utenkelig uten et komplekst, dedikert proteinapparat. Dermed presenterer oversettelsessystemet oss med "Darwin-Eigen-paradokset" som er iboende for alle å tenke på fremveksten av komplekse biologiske enheter: for en moderne type, effektiv og nøyaktig.
Oversettelsessystem For å fungere, er det nødvendig med mange forskjellige proteiner, men for at proteiner skal utvikle seg, ville et oversettelsessystem nesten like bra som det moderne ville være nødvendig.
 RNA -syntese krever RNA -reparasjonsenzymer
RNA -syntese krever RNA -reparasjonsenzymer
En celle har en stor investering i RNA -ene - de jobber kopier av dens genomiske informasjon. Studien av mRNA -biogenese de siste årene har avdekket en forseggjort overvåkningsmekanisme som involverer faktorer som UPF -proteiner som kaster seg avvikende, spleiset mRNA og mRNA med for tidlig terminerings-kodoner. Det kan være et hint om at slike RNA -kvalitetskontrollmekanismer går galt i kreftformer, akkurat som DNA -kvalitetskontrollmekanismer gjør, der aberrant spleisede transkripsjoner akkumuleres i en svulst. Nå som portene er åpne, kan vi ha en flom av studier på RNOME [RNA -genomet] stabilitet og kreft.
Bilde 7. Litt ironi må påregnes
Dette forverrer kylling-og-egg-problemet for talsmenn for naturlige mekanismer. I "RNA -verden" -hypotesen for livets opprinnelse, utførte RNA både informasjonslagring og enzymatiske funksjoner før disse rollene ble outsourcet til DNA og proteiner. Men hvordan kunne RNA reparere seg selv? Hvis RNA må beskyttes mot skade, ville proteinreparasjonssystemet ha trengt å være der fra begynnelsen. Talsmenn for naturlige mekanismer kan anta at forskjellige primitive RNA jobbet side om side for å reparere hverandre, men at det anstrenger troverdighet for en hypotese allerede er langsiktig. I typisk evolusjonær lingo røyklegger Begley og Samson hva i naturen som ble produsert: "Det ser ut til at parametere for hvert menneskelig protein har utviklet seg for å skille mellom RNA og DNA," spekulerer de og et annet sted, "det Kan være at RNA-demetyleringsaktiviteten til alkb-lignende proteiner utviklet seg for å regulere biologisk RNA-metylering, og at reparasjon av avvikende, kjemisk metylering er heldig. ” Spør dem hvordan cellen utviklet disse tingene, og du vil sannsynligvis få et spørrende utseende, som om "hvorfor stiller du et så stumt spørsmål? Jeg vet ikke. Det måtte bare. Vi er her, ikke sant? ” (7)
Du trenger energi for å lage energi
Når Jorden hadde pyrofosfitt, hadde den et energisk molekyl som, selv om den ikke var så nyttig som ATP, i det minste var noe likt. "Teamet fant at en forbindelse kjent som pyrofosfitt kan ha vært en viktig energikilde for primitive livsformer." Hadde han noen bevis for dette? Nei; Det er bare et krav. "Det er et kylling- og eggspørsmål," sa han. Forskere er uenig om hva som kom først - replikasjon eller metabolisme. Men det er en tredje del av ligningen - og det er energi. ” Så mens forskere er uenige om to ting, hvorfor ikke legge til en tredje? Det er fremgang: "Du trenger enzymer for å lage ATP, og du trenger ATP for å lage enzymer," forklarte Dr. Kee, som om å legge til spørsmål kvalifiserer som å forklare noe: "Spørsmålet er: hvor kom energi fra før en av disse to tingene eksisterte? ” Vi vet kanskje ikke svarene, men i det minste blir uvitenheten vår mer sofistikert takket være OTL -forskning. (17)
Maskinvaren og programvaren til cellen, bevis på design (18=
Paul Davies: The Fifth Miracle Side 62
På grunn av organisasjonsstrukturen til systemer som er i stand til å behandle algoritmisk (instruksjons) informasjon, er det overhodet ikke klart at et monomolekylært system - der en enkelt polymer spiller rollen som katalysator og informasjonsbærer - er til og med logisk konsistent med organiseringen av informasjonsstrømmen i levende systemer, fordi det ikke er noen mulighet for å skille informasjonslagring fra informasjonsbehandling (det å være et så særegent trekk ved det moderne liv). Som sådan representerer digitale første systemer (som for øyeblikket posert) en ganske triviell form for informasjonsbehandling som ikke klarer å fange opp den logiske strukturen i livet slik vi kjenner den. (1)
Vi må forklare opprinnelsen til både maskinvare- og programvareaspektene ved livet, eller jobben er bare halvparten ferdig. å forklare det kjemiske underlaget i livet og hevde det som en løsning på livets opprinnelse er som å peke på silisium og kobber som en forklaring på gang på en datamaskin. Det er denne overgangen der man bør forvente å se et kjemisk system bokstavelig talt ta på seg "et eget liv", preget av informasjonsdynamikk som blir frakoblet fra dikterene til lokal kjemi alene (mens de selvfølgelig forblir fullt konsistent med disse diktene) . Dermed er det berømte kylling-eller-egg-problemet (et utelukkende maskinvarespørsmål) ikke det sanne stikkpunktet. Snarere ligger puslespillet med noe fundamentalt annerledes, et problem med årsaksorganisasjon som har å gjøre med separasjon av informasjonsmessige og mekaniske aspekter i parallelle årsaksfortellinger. Den virkelige utfordringen med livets opprinnelse er således å forklare hvordan instruksjonsinformasjons-kontrollsystemer dukker opp naturlig og spontant fra bare molekylær dynamikk.
Programvare og maskinvare er irreducible kompliserte og gjensidig avhengige. Det er ingen grunn til at informasjonsbehandlingsmaskiner eksisterer uten programvaren, og omvendt.
Systemer for sammenkoblet programvare og maskinvare er irreducible kompliserte.
Bilde 8. Kylling egg problem -ang hjerte og blodkar
 Alle cellulære funksjoner er irreducible kompliserte
Alle cellulære funksjoner er irreducible kompliserte
Paul Davies, The Fifth Miracle Side 53:
Plukk DNA fra en levende celle, og den ville være strandet, ikke i stand til å utføre sin kjente rolle. Bare innenfor konteksten av et meget spesifikt molekylært miljø vil et gitt molekyl spille sin rolle i livet. For å fungere ordentlig, må DNA være en del av et stort team, med hvert molekyl som utfører sin tildelte oppgave sammen med de andre på en samarbeidsvillig måte. å anerkjenne gjensidig avhengig av komponentmolekylene i en levende organisme presenterer oss umiddelbart et sterkt filosofisk puslespill. Hvis alt trenger alt annet, hvordan oppstod fellesskapet med molekyler noen gang i utgangspunktet? Siden de fleste store molekyler som trengs for livet, bare produseres av levende organismer, og ikke finnes utenfor cellen, hvordan kom de til å eksistere opprinnelig, uten hjelp fra en ekstern forsker? Kan vi seriøst forvente at en Miller-Oury-type suppe vil gjøre dem alle på en gang, gitt den hit-and-miss naturen til dens kjemi? (19)
Å være en del av et stort team, samarbeidende måte, inter -pålitelighet, alt trenger alt annet, er bare andre ord for irreducibilitet og gjensidig avhengighet.
For et ikke-levende system er spørsmål om irreducibel kompleksitet enda mer utfordrende for et helt naturlig scenario som ikke er designet, fordi naturlig seleksjon-som er hovedmekanismen for darwinistisk evolusjon-ikke kan eksistere før et system kan reprodusere seg. For livets opprinnelse kan vi tenke på den minimale kompleksiteten som vil være nødvendig for reproduksjon og andre grunnleggende livsfunksjoner. De fleste forskere tror dette vil kreve hundrevis av biomolekylære deler, ikke bare de fem delene i en enkel musefelt eller i mitt imaginære LMNOP -system. Og nåværende vitenskap har ingen sannsynlige teorier for å forklare hvordan den minimale kompleksiteten som kreves for livet (og begynnelsen av biologisk naturlig seleksjon) kunne ha blitt produsert ved naturlige prosesser før begynnelsen av biologisk naturlig seleksjon.
Mens hjertet fungerer som en "pumpe" for å levere blod i hele kroppen, krever det også oksygenrikt blod for å forbli sunt. Eller vurder at produksjonen av røde blodlegemer, en av de viktigste bestanddelene i det vaskulære systemet, er regulert av erytropoietin - et hormon produsert i nyren. Likevel krever den nyren røde blodlegemer for å levere oksygenrikt blod. Så hva utviklet seg først? (22)
Gjensidig avhengighet av lipidmembraner og membranproteiner
Selv i de enkleste cellene er membranen en biologisk enhet av en svimlende kompleksitet som bærer forskjellige proteinkomplekser som medierer energiavhengig - og tett regulert - import og eksport av metabolitter og polymerer
Bemerkelsesverdig, til og med forfatteren av boken: Agents Under Fire: Materialism and the Rationality of Science, PGS. 104-105 (Rowman & Littlefield, 2004). HT: Env. Spør leserne: Derfor ville et kylling- og eggparadoks: En lipidmembran ville være ubrukelig uten membranproteiner, men hvordan kunne membranproteiner ha utviklet seg i fravær av funksjonelle membraner?
 Biosyntese av cellemembraner: Det tar membraner å lage membraner
Biosyntese av cellemembraner: Det tar membraner å lage membraner
Cellevegger gir faktisk essensiell strukturell støtte og ytre interaksjoner i moderne organismer (Albers & Meyer, 2011), til tross for det fantastiske mangfoldet som eksisterer blant prokaryote cellekonvolutter, syntese av mange av deres viktigste komponenter:
..
En er avhengig av sammenlignbare glykosyleringsveier Disse traséene er alle lokalisert i cellemembranene, er mediert av lignende lipidbærere og har samme orientering over membranen.
Det reiser spørsmålet: Hvis i eksisterende celler er disse traséene plassert i cellemembranene, hvor skulle en plassere det foregikk tidligere membraner? Ville det ikke være riktig å si: Det krever membraner å lage membraner?
Referanser:
1) https://reasonandscience.catsboard.com/t2117-nuclear-pores#3762
2) http://www.evolutionnews.org/2011/08/direct_rna_templating_a_failed050121.html
3) http://www.fis.puc.cl/~jalfaro/astrobiologia/Astrobiologiavasquez.pdf
4) Meyer, signature of the cell, page 111
5) https://reasonandscience.catsboard.com/t2285-iron-sulfur-clusters-basic-building-blocks-for-life#4646
6) https://reasonandscience.catsboard.com/t2043-dna-error-checking-and-repair#4669
7) https://reasonandscience.catsboard.com/t2043-dna-and-rna-error-checking-and-repair-amazing-evidence-of-design#4671
8 https://reasonandscience.catsboard.com/t1849p30-dna-replication-of-prokaryotes#4672
9) http://www.ideacenter.org/contentmgr/showdetails.php/id/845
10) https://reasonandscience.catsboard.com/t1796-glycolysis?highlight=glycolysis
11) http://www.scientificamerican.com/article/a-simpler-origin-for-life/
12) http://www.uncommondescent.com/intelligent-design/do-viruses-help-explain-the-origin-of-life/
13) https://en.wikipedia.org/wiki/Reverse_transcriptase#In_eukaryotes
14) Fazale Rana, Cell's design, page 186
16) https://reasonandscience.catsboard.com/t1331-the-cell-membrane-irreducible-complexity
17) http://creationsafaris.com/crev201005.htm
18) https://reasonandscience.catsboard.com/t2221-the-hardware-and-software-of-the-cell-evidence-of-design?highlight=software
19) https://reasonandscience.catsboard.com/t2179-the-cell-is-a-interdependent-irreducible-complex-system
20) https://reasonandscience.catsboard.com/t2285-iron-sulfur-clusters-basic-building-blocks-for-life
21) https://reasonandscience.catsboard.com/t2028-origin-of-the-dna-double-helix#3426
22) http://www.apologeticspress.org/ApPubPage.aspx?pub=1&issue=571&article=450
23) https://reasonandscience.catsboard.com/t2371-how-cellular-enzymatic-and-metabolic-networks-point-to-design#5139
24) https://www.quantamagazine.org/lifes-first-molecule-was-protein-not-rna-new-model-suggests-20171102/
Oversettelse via google oversetter og bilder ved Asbjørn E. Lund